
Рис. 7. Ствол дерева, покрытый эпифитами (тропический, вечиозеленый, сырой лес Бразилии).
| Сожительство и растительныя сообщества |
|
Е. Варминг |
Между растениями существуют разнообразныя связи неодинаковой прочности. В одних случаях сожительство известных видов влияет глубоко на жизнь каждаго из них, в других случаях существующая между видами связь менее тесна или даже совсем случайна. Мы начнем наше изложение с таких форм сожительства, где между видами существует самая тесная, а именно органическая связь (симбиоз в широком смысле слова); далее мы разсмотрим формы сожительства менее прочныя и, наконец, остановимся более подробно на наименее тесно связанных формах сожительства, обнимающих собой большое число форм. Все эти формы сожительства не резко разграничены между собой.
Паразитизм - это такая форма сожительства, где между обоими живущими совместно видами существует наиболее тесная связь. Один вид снабжает пищей другой, паразит живет на своем "хозяине", или в нем, на счет его живых тканей. Существуют, однако, различия в том, как паразит связан с хозяином. Наиболее зависят от хозяев ржавчинные грибы и льняная павилика (Cuscuta Epilinum) или виды Orobanche и т. д., являющиеся не только полными паразитами, т. е. не могущими питаться неорганической пищей, но и приуроченными только к определенным видам растений.
В меньшей зависимости от своего хозяина находятся виды растений, могущие произрастать на нескольких или даже на многих видах растений, принадлежащих к одному или даже к различным семействам, например, Cuscuta Epithymum (полный паразит) может жить на Calluna, Labiatae, Papilionacoae и др., a Viscum album (полупаразитный вид) на полусотне видов лиственных и хвойных деревьев.
В то время, как одни виды, обязательные паразиты (облигатные), могут вести только паразитный образ жизни, другие виды не так стеснены в этом отношении и в некоторых случаях могут существовать, как сапрофиты (растения, живущия на гниющих веществах), например, опенок (Armillaria mellea), Nectria cinnabarina и др.
Между паразитом и его хозяином существуют враждебныя (одностороннеантагонистическия) отношения: паразит истощает своего хозяина. Истощение хозяина может быть так велико, что паразит убивает его (Loranthaceae в состоянии, например, лишить жизни померанцевое дерево); конечно, в таком случае паразит часто также погибает.
Борьба между данным видом и его паразитом имеет большое значение для состава растительных сообществ. Многия лесныя деревья погибают в борьбе с грибами, что оказывает влияние на лесную растительность целой страны, например, в Дании. Разводимые леса подвергаются в большей степени нападению грибов, чем туземные, что обусловливается тем, что в однообразном насаждении паразиты могут распространяться с большей легкостью. Нападение паразитов, наряду с климатическими условиями, является часто причиной вытеснения одного вида растений другим.
Гелотизм. Сожительство лишайников с водорослями лучше всего обозначать названием гелотизм. Лишайник - двойной организм, состоящий из гриба и водоросли. Эта последняя окружается гифами гриба и заключается в его ткани. Отношение это считается обыкновенно мутуалистическим, т. е. предполагают, что оба организма оказывают друг другу известныя услуги, и это до некоторой степени верно: водоросль, благодаря своему хлорофиллу, снабжает гриб углеродистыми веществами и содействует переработке пищи, между тем как на долю гриба приходится все остальное. Связь, однако, не равная для обеих сторон: гриб необходимо должен соединяться с водорослью для своего полнаго развития, но водоросль не нуждается в грибе и, по всей вероятности, вне его произрастает лучше. Поэтому-то название "Konsortium" тоже не совсем подходить для этого рода сожительства. Причина сильнаго роста и быстраго размножения водоросли, клетки которой бывают в этих условиях больше, чем когда водоросль развивается вне гриба, не что иное, как гипертрофия, болезненное состояние растения. Предполагали, что водоросль находить в теле гриба защиту против высыхания. Но предположение это неверно, так как водоросль в этом вовсе не нуждается и прекрасно переносит всякое высыхание, и, кроме того, в известных условиях лишайник до того высыхает, что делается хрупким. Высыхание препятствует, конечно, более совершенному способу размножения водоросли, а именно посредством зооспор. Очевидно, водоросль находится в грибе как бы в рабстве и гриб является своего рода паразитом, отличающимся от обыкновеннаго паразита тем, что он заключает в себе своего хозяина и добывает часть своей пищи. Очевидно, тут существует некоторое сходство с зелеными полупаразитами, с той лишь разницей, что эти последние сами добывают углеродистую пищу, между тем как лишайниковый гриб заботится исключительно о не содержащей углерода пище.
И тут также связь между обоими организмами может быть весьма тесной, причем гриб может жить лишь с известным видом водорослей.
Сомнительно, существует ли вообще среди растений обоюдный мутуализи, где бы сожительство было одинаково полезно для обеих сторон. Большая часть условий сожительства организмов недостаточно нам известна для того, чтобы дать себе полный отчет о существующей между ними связи. Это относится, например, к Mykorrhiza, где корни высших растений вступают в тесную, экто - или эндотрофическую связь с безплодными гифами гриба, т. е. или с гифами, образующими колпачок на поверхности корневых кончиков, или же с гифами, живущими в клетках коры корней. Mykorrhiza найдена у большинства сережчатых, хвойных, Ericaceae и некоторых других, преимущественно многолетних травянистых растений, живущих на кислом перегное, на торфянистой и перегнойной почве, т. е. вообще на богатых гумусом почвах. Весьма возможно, что мицелии извлекают известную пользу из цветковых растений и, почти несомненно, приносят пользу этим последним. Внекоторых случаях они замещают корневые волоски и, по всей вероятности, извлекают из богатой гумусом почвы органическую, содержащую азот пищу. Если это предположение, относящееся, главным образом, к эктотрофическим Mykorrhiza, верно, то мы имеем здесь замечательный пример того, что одине вид растения может оказывать содействие другому при заселении и добывании пищи в таких местах, который иначе были бы недоступны для даннаго вида растений. Верещатники и еловые леса обязаны, таким образом, своим существованием до некоторой степени этой форме сожительства.
На микорицу, особенно эндотрофическую, походят до некоторой степени бактерии, сожительствующия с бобовыми. Бактерии, проникающия извне в корни бобовых, находят в них помещение и вызывают в них образование клубеньков; оне усвояют пищу, содержащую азот, затем погибают, превращаясь в "бактероиды", и, наконец, идут на пищу растению. С достоверностью неизвестно, извлекают ли бактерии пользу из этого сожительства (по всей вероятности, оне получают от хозяина углеродистую пищу), но, с другой стороны, было бы очень странно, если бы оне проникали в корни, как эндотрофические грибы, без всякой для себя пользы.
У Alnus, Elaeagnaceae, Myrica и Ceanothus также встречаются такие же корневые клубеньки, образованные однако, не бактериями, а гифами гриба Frankia.
Идя дальше, мы придем к растениям (водорослям), пользующимся другими растениями, как помещением, но не оказывающим им, насколько это известно, взаимной пользы. Они не живут на счет своего хозяина и ничего, быть может, из него не берут, но живут до некоторой степени свободно. Сюда принадлежит одна сине-зеленая водоросль (Anabaena), живущая в особых полостях на нижней стороне листьев Azolla; полости эти, кажется, специально приспособлены для нея, постоянно находятся у всех четырех видов Azolla и никогда не бывают свободны от Anabaena. Водоросль может существовать вполне хорошо и вне Azolla. Точно также эндофитно, т. е. внутри других растений, живут и некоторыя другия водоросли; например, в листья Sphagnum проникают через отверстия в безцветных клетках нити Nostoc; в некоторых печеночных мхах или в других водорослях же тоже поселяются водоросли, например, Entoderma viride в клеточной оболочке Derbesia Lamourouxii.Может быть, однако, что в последнем случае мы имеем дело с паразитизмом.
Отчасти это происходить, вероятно, также с Cyanophyсеае, проникающими в вертикальные дихотомически разветвленные корни Cycadeae, причем они заставляют известныя клетки паренхимы разрастаться специальным образом и этим самым готовят себе в них помещение. Главным же образом это происходит с водорослями (Nostoc), проникающими в стволы Gunnera, но могущими прекрасно существовать и вне этих стволов (ср. Jonsson в Bot. Notiser, 1894). Настоящия наши познания не дают еще нам возможности получить ясное представление о природе этой формы сожительства.
Эпифиты. От эндофитов, находящих лишь помещение внутри других растений, существует незаметный переход к растениям, живущим эпифитно, т. е. на поверхности других видов растений. Они не извлекают пищи из живых тканей этих последних и в крайнем лишь случае питаются их мертвыми тканями. Однако, не всегда можно с уверенностью сказать, что они не живут на счет своего хозяина, так как эпифиты могут появляться на других растениях в таком количестве, что приходится допустить, что они приносят вред своим количеством, вызывая слишком большую влажность или затрудняя дыхание этих последних (рис. 7, стр. 124).
Связь между эпифитами и видами, на которых они живут, обыкновенно менее тесная, чем в предыдущем случае; многие эпифиты могут существовать на многих видах растений, а некоторые также и на скалах. Некоторые из них, однако, связаны с определенными видами, так как для них природа коры имеет известное значение. Эпифиты в?тречаются на сухопутных и на водных растениях. Многия водоросли живут на других водорослях или на цветковых растениях, но некоторыя из них только на определенныхвидах, например, Elachista fucicola на Fucus, E. scutulata на Himanthalia lorea и т. д.
На сухопутных растениях эпифиты произрастают с наибольшим успехом во влажном воздухе и в местах, изобилующих осадками. На это обратил внимание уже Meyen, позднее Schimper в своей работе об эпифитах (I, III) разработал этот вопрос более обстоятельно. В холодных и умеренных странах эпифитами являются, главным образом, водоросли, лишайники и мхи, но в жарких странах к ним присоединяются также многочисленные папоротники и цветковыя растения, принадлежащия к различным семействам (Orchidaceae, Araceae, Bromeliaсеае, Рирегасеае и др.). В тропических сырых лесах сюда присоединяются еще эпифиллы, т. е. виды, живущие на многолетних листьях других растений (см. 6 отд., гл. VII).
Особенности местонахождений вызвали некоторыя биологическия приспособления, значение которых, по Schimper'y для цветковых растений состоит в следующем.
Семена (и споры) обладают двумя родами приспособлений, служащих для их переноса и укрепления на субстрате. В одних случаях они незначительных размеров, легки, снабжены длинными волосками, благодаря чему легко уносятся ветром и попадают на стволы и ветви, где и укрепляются в трещинах или углублениях, или же они бывают заключены в сочные плоды, которые служат пищей птицам, после чего семена распространяются вместе с экскрементами и укрепляются на ветвях (Araceae, Bromeliaceae, Cacteae). Лишенная корней Tillandsia usneoides распространяется своеобразным образом: оторвавшиеся куски ея длинных, тонких побегов легко обвиваются вокруг ветвей деревьев (рис. 8, стр. 126).
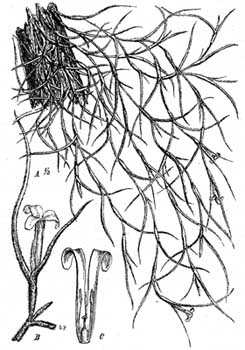
Укрепление эпифитов на растениях происходит или посредством ризоидов, проникающих не глубоко в субстрат (отмершия части коры), например, у мхов, лишайников, или посредством цепких корней, очень раздражимых и прикрепляющихся к субстрату посредством присасывающихся волосков. Между прикрепляющимися корнями (присосками) и всасывающими воду корнями существует известное разделение труда.
Водоснабжение составляет для эпифитов трудную задачу, так как дождевая вода скоро стекает. По всей вероятности, они получают больше воды из росы и тумана, чем из дождя. Многие приспособлены к тому, чтобы пользоваться каждой благоприятной минутой, и могут в сухом виде поглощать влагу всей своей поверхностью (водоросли мхи, лишайники и Tillandsia usneoides, снабженная, как и другия Bromeliaceae, особенными всасывающими волосками). Другие виды (Orchidaceae, Araceae) имеют воздушные корни, снабженные особой корневой оболочкой, приспособленной для восприятия воды; еще другие, например, Tillandsia bulbosa, имеют листья такого устройства, что вода может задерживаться между ними, и, наконец, есть еще растения, имеющия два рода листьев, из которых одни плотно прижимаются к стволам, благодаря чему в состоянии капиллярно удерживать воду между, собой и поверхностью ствола; быть может, они также в состоянии поглощать воду (например, папоротник Teratophyllum aculeatum по G. Karsten'y). Эпифиты часто подвергаются высыханию. Против этого некоторые из них не обладают никакими очевидными приспособлениями (водоросли, лишайники, мхи); они могут переносить без вреда для себя продолжительную засуху и при первом дожде или росе опять пробуждаются к жизни. Другие виды выработали у себя водовместилища разнаго устройства: водоносную ткань в стеблях и листьях (Orchidaceae, Peperomiaceae и др. рис. 2); водоносныя, клетки в листьях (Orchidaceae, рис. 9); кувшинообразныя и другия углубления (печеночные мхи по Goebel'ю; Dischidia и др., рис. 10, стр. 128).
Пищу эпифиты добывают следующим образом: углерод они черпают из воздуха, т. к. принадлежат всегда к живущим на свету и вечнозеленым растениям; некоторые собирают, кроме того, между корнями или помощью особенных листьев гумусовыя и минеральныя частицы (листья с углублениями или в виде мантий), например некоторые папоротники (Asplenium Nidus, Polypodium quercifolium, Platycerium alcicorne; Goebel, II, 1-ая часть). Рис. 11, стр. 129. Строение побегов, а также все устройство эпифитов могут быть весьма различны. Существуют виды, лишенные корней (Tillandsia usneoides), и виды, вегетативные органы которых состоят почти всецело из зеленых корней, например, Polyrrhiza funalis (Acranthus funalis; Orchidaceae). Следуя Schimper'y, можно разделить эпифиты на четыре группы: 1) на эпифиты, получающие всегда пищу из коры, к которой они прикреплены; 2) на виды, пускающие в землю воздушные корни; 3) на эпифиты, воздушные корни которых образуют густое сплетение, где накопляется влажный гумус; 4) наконец, эпифиты, у которых листья исполняют деятельность корней и поглощают влагу и питательныя соли (ср. также Karsten, III). Эпифиты имеют в своем строении много общаго с растущими на земле ксерофитами, потому что они, подобно последним, приспособлены к продолжительной засухе: они составляют, в сущности, лишь группу ксерофитов, поэтому-то понятно, почему некоторые виды могут жить не только на деревьях, но и на скалах (например, Rhipsalis Cassytha и другия кактусовыя). Странныя особенности их строения будут разобраны в 4-м отделе (ксерофитная растительность).
Сапрофиты (перегнойныя растения). Мы должны допустить, что многие эпифиты извлекают пищу из мертвых растительных частей (например, из коры), на которых они растут. Стало быть, они питаются мертвыми органическими веществами, т. е. сапрофиты.

Большее количество сапрофитов и более характерныя формы их попадаются, однако, прямо на земле, в особенности в лесах, где из года в год накопляются всякаго рода отбросы (опавшие листья, ветви, цветки и плоды), образующие гумус.
Сапрофиты, стало быть, также связаны с другими разным образом, дети лесов и кустарников (рис. 12). Зaтемнение. причиняемое густой растительностью, заставило их первоначально вытягиваться кверху, образовать длинночленистые побеги и постепенно приспособляться к тому, чтобы удержаться в таком положении, а, вместе с тем, относительно внутренняго строения решить трудную задачу перемещения веществ по весьма длинным и тонким стеблям (подробнее см. Schenck, VI, Warming, VIII). Строение листа и побегов у некорых лиан напоминает строение ксерофитов, что, конечно, весьма естественно, так как лианы могут терять путем испарения много воды и эта потеря не всегда может быть вполне врзмещена деятельностью корней; строение их должно быть, конечно, приспособлено к этому условию жизни (Warming, VIII). Форма лиан вызвана условиями общественной жизни, но, с другой стороны, лианы отчасти независимы от других растений, так как в некоторых случаях мертвыя подставки могут им служить опорой вместо живых.
В этой главе мы разсмотрели разнообразные случаи связи между растениями. Прежде всего мы остановились
на связи между отдельными неделимыми: между паразитом и хозяином, между господином и рабом (гелотизм
лишайников); дальше мы разсмотрели мутуалистов и эпифитов и перешли, наконец, к видам, связанным с
целыми растительными обществами. Нам осталось еще разсмотреть большия, очень сложныя растительный
сообщества, составляющия собственно предмет ойкологической географии растений.